 Kelly lab. Mike, Mark, Craig, Dave, Gavin, Marie, Steve, Stu, John & Jyoti.
Kelly lab. Mike, Mark, Craig, Dave, Gavin, Marie, Steve, Stu, John & Jyoti.
The tripartite ATP-independent periplasmic (TRAP) transporter family has been a major interest of the Thomas lab since it was founded in 2002 in York. Gavin had worked with Prof. David Kelly at the University of Sheffield from 2000 as a postdoc on a BBSRC funded grant to characterise the functional diversity of TRAP transporters, which had been discovered 5 years earlier in Dave's lab. (Wyborn et al., 1997, J. Bact.) Around the same time Arnold Driessen's & Wil Koning's group in Groningen, Netherlands, described a binding-protein-dependent secondary transporter, which was almost undoubtedly a TRAP transporter also (Jacobs et al., 1996). The hook here then is that these are bacterial transporters that use the 'high affinity' properties of a substrate binding protein (SBP) but instead of coupling this to ATP binding and hydrolysis, like in an ABC transporter, they use the membrane potential - a more efficient energy source - to drive active transport. We set out to develop a model transporter to probe how TRAP transporters work.
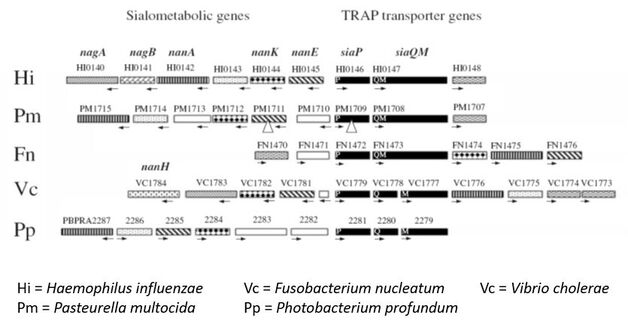 Genome gazing led to the identification of the sialic acid TRAP transporter, SiaPQM.
Genome gazing led to the identification of the sialic acid TRAP transporter, SiaPQM.
Early days - 2000-2005 When working in the Kelly lab, one of my first jobs was to pull together the known scattered literature on TRAP transporters (Kelly & Thomas, 2001), and in the process of doing this we did a deep mine of the then current bacterial genome sequences to look for potentially novel and interesting systems (this was the early days of genomics so there weren't that many). Initially we worked on Escherichia coli as this has only a single TRAP transporter, but while we were able to show that this was a 2,3-diketogulonate transporter, its function didn't seem critical to E. coli (Thomas et al., 2006). Gavin found one system on the genome of the pathogen Haemophilus influenzae, which was sitting in an cluster of genes looked like they were involved in sialic acid catabolism. Many were similar to those discovered by Eric Vimr at the University of Illinois, USA, that enabled Escherichia coli to use sialic acid as the sole carbon and energy source. The E. coli nan cluster, as it was known, contains an MFS transporter, NanT, but this was seemingly replaced in H. influenzae by the putative TRAP transporter. The known substrates for TRAP transporter were C4-dicarboxylates, such as the citric acid cycle intermediate, fumarate and succinate. Sialic acid was also an organic acid, so seemed like it could be a 'TRAP substrate'. Hence, this system become our primary study system for TRAP transporters.
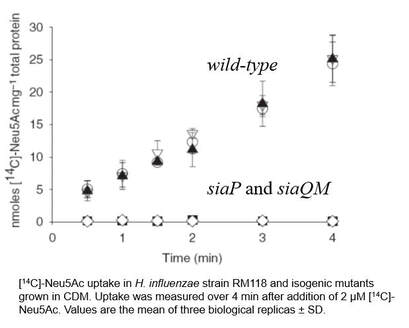
First grant - Our first grant from BBSRC started in 2005, which enabled Dr. Emmanuele Severi to join the group and characterise the H. influenzae TRAP transporter experimentally, which we named SiaPQM, with SiaP encoding the soluble substrate binding protein (SBP) and SiaQ and SiaM being the small and large membrane subunits, although neatly in this case they were fused into a single protein, SiaQM.
Emm showed that these genes were required for growth on sialic acid, which was through a direct function in uptake (see figure). Then working with Derek Hood's group in Cambridge, we were able to show that siaPQM is important for survival of H. influenzae in human serum (Severi et al., 2005). Emm also did some great work discovering other sialic acid transporters and novel catabolic enzymes (see the Sialic acid biology research page).
Emm showed that these genes were required for growth on sialic acid, which was through a direct function in uptake (see figure). Then working with Derek Hood's group in Cambridge, we were able to show that siaPQM is important for survival of H. influenzae in human serum (Severi et al., 2005). Emm also did some great work discovering other sialic acid transporters and novel catabolic enzymes (see the Sialic acid biology research page).
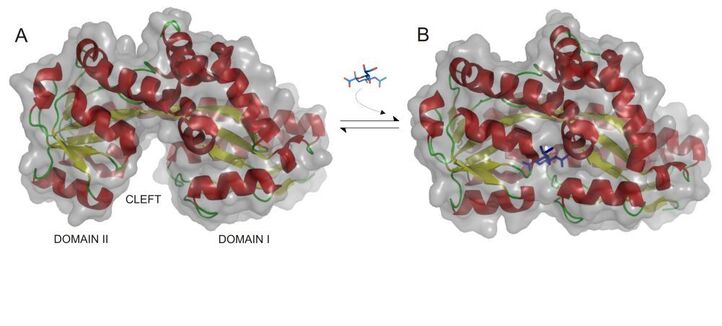 The structure of SiaP in the open unliganded and close liganded forms. The Venus flytrap movement.
The structure of SiaP in the open unliganded and close liganded forms. The Venus flytrap movement.
A new PhD student, Chris Mulligan, joined the group shortly afterwards, who started the painstaking work on the membrane domains themselves, first working on the sole E. coli TRAP transporter , but then soon after on SiaQM. Around this time we also started a long-term and highly productive collaboration with Prof. Anthony Wilkinson and Keith Wilson in the York Structural Biology Laboratory which was just along the corridor from the lab. We persuaded a brilliant PhD student Alex Muller to work on SiaP as part of his project and he solved the first ever structure of an SBP from a TRAP transporter, which was super cool and revealed the molecular details of the first interaction in the transport cycle between the ligand (sialic acid in this case) and the transporter (Muller et al., 2006). Shortly after a structure of SiaP from another strain of H. influenzae was solved with the sialic acid (Neu5Ac) bound, rather than the analogue we had in our structure. This was super useful as now there are structures of over 100 different TRAP SBPs bound to, predominantly, organic anions.
Our first structure and bioinformatics also revealed a key conserved arginine residue in SiaP which interacts with the carboxylate group on the ligand. This is a key feature of most TRAP SBPs and relates to the ligand range of TRAP transporters, which we'll come back to later. Chris had also done much deeper mining of the hundreds of newle sequenced bacterial genomes, and we found that TRAP transporter where enriched in marine bacteria, and hence we proposed that they might be using Na+ ions rather than H+ as the coupling ion for transport (Mulligan et al., 2007).
Our first structure and bioinformatics also revealed a key conserved arginine residue in SiaP which interacts with the carboxylate group on the ligand. This is a key feature of most TRAP SBPs and relates to the ligand range of TRAP transporters, which we'll come back to later. Chris had also done much deeper mining of the hundreds of newle sequenced bacterial genomes, and we found that TRAP transporter where enriched in marine bacteria, and hence we proposed that they might be using Na+ ions rather than H+ as the coupling ion for transport (Mulligan et al., 2007).
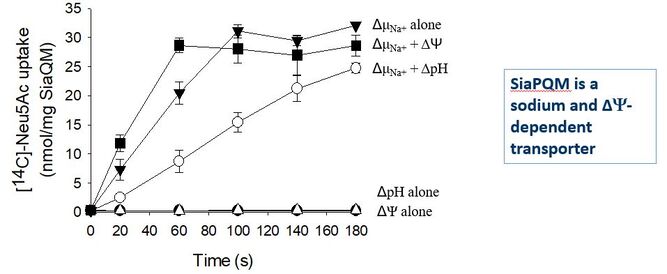
Going forward with the second grant.
Towards the end of Chris's PhD project, after limited success using the E. coli TRAP systems, we teamed up with Bert Poolman's team in Groningen. Chris worked with Eric Geertsma to get enough protein for reconstitution and get our first direct measurements of function. In some beautiful work they were able to fully reconstitute SiaPQM function and we were able to show that the system absolutely needed the SBP, is Na+ dependent (as we had proposed) and was electrogenic (i.e. as the sialic acid has a net negative charge, at least two positive charges are needed to drive its concentrative uptake into the cell) (Mulligan, Geertsma, et al., 2009)
With Fidelma Boyd the University of Cork, Ireland, and the University of Delaware, USA, we studied the related system in the human pathogen V. cholerae, which had similar properties to the H. influenzae system and was also involved in colonisation, although just as a nutrient source and the bacterium does not sialylyate its LPS like H. influenzae (Chowdhury et al., 2012). This also involved some work showing that a TRAP transporter, that another group had claimed was a sialic acid transporter, was in fact specific for C4-dicarboxylates (Thomas & Boyd, 2011).
With this new methodological breakthrough we obtained more support from BBSRC in 2008, enabling Chris to continue his work on the membrane domains. We then switched to the V. cholerae system, which was truly tripartite, and Chris showed that Q and M form a 1:1 complex in the membrane (Mulligan et al., 2012). With this information we could then think more about the SBP again and how it functions as a key part of the transporter.
Towards the end of Chris's PhD project, after limited success using the E. coli TRAP systems, we teamed up with Bert Poolman's team in Groningen. Chris worked with Eric Geertsma to get enough protein for reconstitution and get our first direct measurements of function. In some beautiful work they were able to fully reconstitute SiaPQM function and we were able to show that the system absolutely needed the SBP, is Na+ dependent (as we had proposed) and was electrogenic (i.e. as the sialic acid has a net negative charge, at least two positive charges are needed to drive its concentrative uptake into the cell) (Mulligan, Geertsma, et al., 2009)
With Fidelma Boyd the University of Cork, Ireland, and the University of Delaware, USA, we studied the related system in the human pathogen V. cholerae, which had similar properties to the H. influenzae system and was also involved in colonisation, although just as a nutrient source and the bacterium does not sialylyate its LPS like H. influenzae (Chowdhury et al., 2012). This also involved some work showing that a TRAP transporter, that another group had claimed was a sialic acid transporter, was in fact specific for C4-dicarboxylates (Thomas & Boyd, 2011).
With this new methodological breakthrough we obtained more support from BBSRC in 2008, enabling Chris to continue his work on the membrane domains. We then switched to the V. cholerae system, which was truly tripartite, and Chris showed that Q and M form a 1:1 complex in the membrane (Mulligan et al., 2012). With this information we could then think more about the SBP again and how it functions as a key part of the transporter.
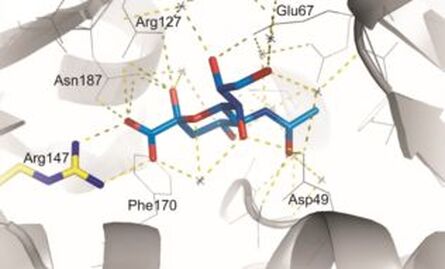
A new PhD student, Adam Hopkins, started to look at the substrate range of SiaPQM and more on the residues in the binding site of SiaP. Importantly he was able to show that SiaPQM can also catalyst uptake of the related sialic acids N-glycolylneuraminic acid (Neu5Gc) and 3-keto-3-deoxy-d-glycero-d-galactonononic acid (KDN), which then got us thinking about how SiaP, which has an enclosed binding site, can accommodate these related ligands (Hopkins, Hawkhead & Thomas, 2013).
About this time we reviewed TRAP transporters 10 years down the line in 2011 (Mulligan, Fischer & Thomas, 2011). Unfortunately our next grant was not funded by BBSRC, Welcome or MRC and so there was a bit of a hiatus in the work and the focus of the group switched to other sugar and peptide transporters (see General Transporters page and the microbiology of human body odour page).
However, some work continued with Marcus Fischer in YSBL where continuing Adam Hopkins's work, another PhD student in the lab, we were able to figure out the role of the conserved arginine (Arg147 in SiaP) which was important for high-affinity and ligand selectivity, but surprisingly not completely essential (Fischer, Hopkins, Severi et al., 2015). Also, working with Rob Hubbard's group in YSBL, Adam had made some mutant we thought might improve the ligand binding specificity of SiaP, but this didn't seem to be the case, but in the end Marcus realised when he crystallised them all that by measuring the thermodynamics of binding we could now determine precisely the contribution of a single water in a water network involved in sialic acid binding. This work took a while to complete and John Darby finished off the experimental work while Marcus, freshly setup with a faculty position at St. Jude Children's Research Hospital, in Tennessee, USA, got his first senior author paper (Darby, Hopkins et al., 2019).
About this time we reviewed TRAP transporters 10 years down the line in 2011 (Mulligan, Fischer & Thomas, 2011). Unfortunately our next grant was not funded by BBSRC, Welcome or MRC and so there was a bit of a hiatus in the work and the focus of the group switched to other sugar and peptide transporters (see General Transporters page and the microbiology of human body odour page).
However, some work continued with Marcus Fischer in YSBL where continuing Adam Hopkins's work, another PhD student in the lab, we were able to figure out the role of the conserved arginine (Arg147 in SiaP) which was important for high-affinity and ligand selectivity, but surprisingly not completely essential (Fischer, Hopkins, Severi et al., 2015). Also, working with Rob Hubbard's group in YSBL, Adam had made some mutant we thought might improve the ligand binding specificity of SiaP, but this didn't seem to be the case, but in the end Marcus realised when he crystallised them all that by measuring the thermodynamics of binding we could now determine precisely the contribution of a single water in a water network involved in sialic acid binding. This work took a while to complete and John Darby finished off the experimental work while Marcus, freshly setup with a faculty position at St. Jude Children's Research Hospital, in Tennessee, USA, got his first senior author paper (Darby, Hopkins et al., 2019).

A new era. After funding ceased on the project, a new collaboration started with Dr. Gregor Hagelueken, who had been a senior postdoc with Jim Naismith FRS at St. Andrew's and was moving back to Germany to the University of Bonn to start his own group. Gregor also thought TRAP transporters were cool and he was able to win funding from the DSF. The groups' research first focussed on using new biophysical tools to analyse the SBP subunit, SiaP, using the V. cholerae system initially. Also, the original TRAP guru, Dr. Emmanuele Severi came back from industry to work on another project in the Thomas lab, but also continued to dabble on the SiaPQM system using tools he had produced almost a decade earlier. The first paper from the collaboration in 2017 was led by Janin Glaenzer who was able to use PELDOR spectroscopy to clearly define two states, open and closed, of SiaP in solution (Glaenzer et al, 2017 Biophys J - with commentary from G Jeschke). Around this time we met Thorbun Cordes, then at Groningen and now in Munich, and figured his single-molecule FRET methods could be really nicely applied to SiaP to examine its resting state and sialic acid-induced closure. Martin Peter and Christian Gebhardt, from Gregor and Thorbun's labs, respectively, used the V. cholerae SiaP to examine this open to closed transition in single molecules of SiaP and finally got our own sialic acid (Neu5Ac) bound forms in the closed state (Peter et al 2021, JMB). As we had suspected and seen from analysis by Marcus Fischer in York (Fischer et al., 2010 Trends Microbiol), the contacts that 'close' the Venus fly trap (open to closed transition) are mediated almost completely by the ligand as the 'glue' that holds the two domains together. Martin continued to work with Christian and Thorbun's group and also produced an important paper compared PELDOR and smFRET methods, using SiaP as a key example (Peter, Gebhardt et al 2022).
The last leg. Martin continued as a postdoc in Gregor's group and worked incredibly hard to develop nanobody/megabody approaches to try and solve the structure by cryo-EM, which he did in the summer of 2022 - ironically at pretty much the same time he also solved the crystal structure of the protein! The structure was a a revelation as it revealed that TRAP transporter are monomeric elevators, in the sense that the small subunit, which we've never really known what it does, acts as a pseudo-stator with the stator domain in the large subunit, such that SiaQM together can function using an elevator mechanism (see figure below). By a nice coincidence, the leading research on a related bacterial elevator protein, the Vibrio cholerae VcINDY protein, had been done by Dr. Chris Mulligan since he last left the lab and now, via NIH, is working at the University of Kent, UK! So, finally after 20 years we actually got to see how TRAP transporters work (Peter et al., 2022), but only through working with exceptional collaborations in the EU.
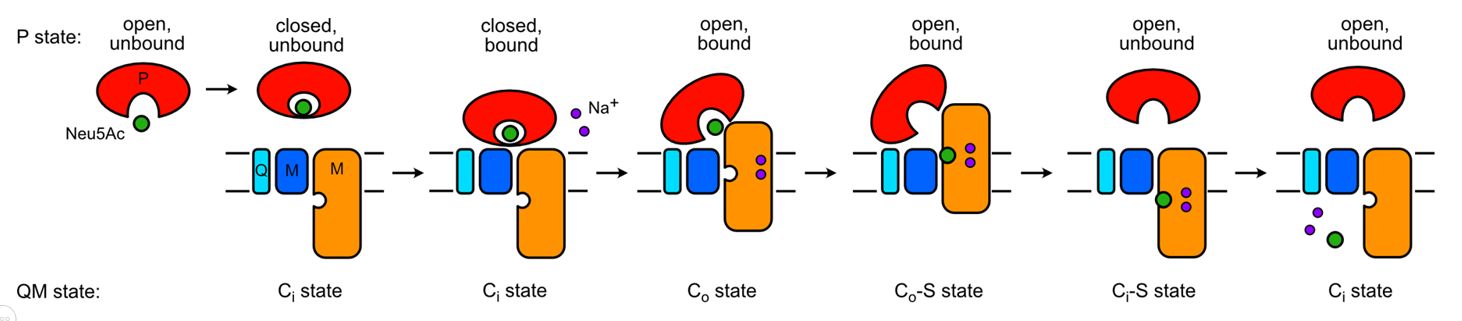
Current working model of the transport cycle of a TRAP transporter (based on Peter et al., 2022).
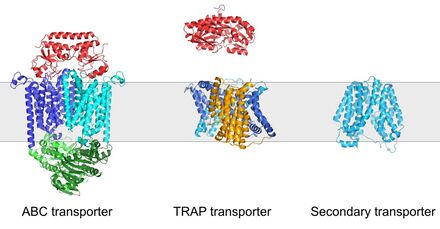 ABC versus TRAP versus MFS. TRAP-Ts contain an SBP like an ABC transporter but use the membrane potential like an MFS transporters. Figure produced by Martin Peter.
ABC versus TRAP versus MFS. TRAP-Ts contain an SBP like an ABC transporter but use the membrane potential like an MFS transporters. Figure produced by Martin Peter.
Going forward. Now we have the structure, the *big* questions can now be asked about the mechanism and there are plenty. How does this monomeric elevator work? Somehow the binding of substrate-loaded SiaP triggers events in the SiaQM protein that 'raise' the elevator to open the SBP, releasing sialic acid into the transporter. This must involve the binding of at least 2 sodium ions, although where and exactly when they bind we don't know. Once the sialic acid is released from the SBP, it must then be 'moved' through the membrane by the 'lowering' of the elevator to then be released on the inside. These are the kind of questions we are now asking of this system. Having ABC and MFS to 'compare and contrast' has been useful in the past, but now we need to look more closely at elevator mechanism-based transporters to fully understand this system, which of course if we could block then would disable H. influenzae and potentially other pathogens like V. cholerae.